Table of Contents
Introduction to Antigen-Presenting Molecules
The immune system defends the host against microbial infections and mutant cells. In defense of the host, the immune system must differentiate between foreign proteins and “self-proteins.” Surface display proteins are used as markers of “self.” Early animal skin transplantation studies demonstrated that rejection or acceptance of grafts was dependent on “self ” markers mapped to a gene cluster on chromosome 17. The major histocompatibility complex was named after this gene cluster (MHC). The human leukocyte antigens (HLA) complex is the name given to the MHC in humans. The word comes from the usage of alloantigens in white blood cells in tissue typing of donors. Before organ transplantation, donors and patients were interviewed. Even though HLAs are crucial in transfusion responses, organ transplantation, and other procedures. An antigen is the most significant role they play in autoimmune and autoimmunity. T cells receive the message. HLA molecules play a key part in this. susceptibility to infection or resistance to infection, and the development of autoimmune diseases antibody-mediated and cell-mediated reactions responses.
The human leukocyte antigen complex
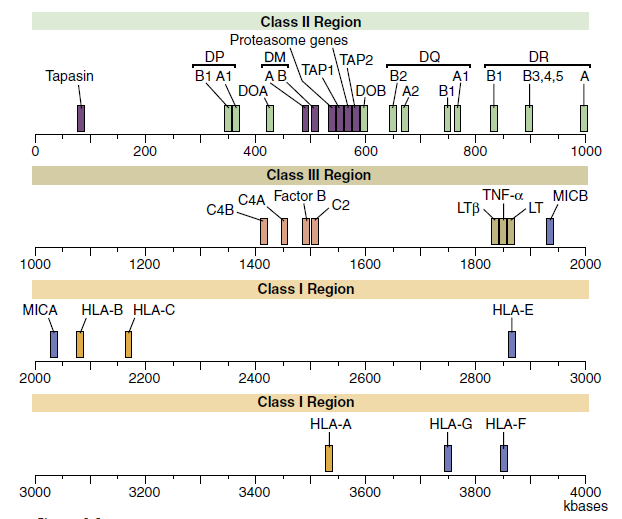
The HLA complex is a gene cluster with 128 functional genes and 96 pseudogenes (partial genes or gene fragments). The majority of gene products are produced in nucleated cells. However, some are secreted proteins that help the body function better. Inflammatory reaction based on the structure of gene products The HLA gene complex is separated into two parts: structure and function. Regions are divided into three categories: class I, class II, and class III. Pseudogenes and their Function Although the role of pseudogenes in coding is unknown, evidence shows that they do. HLA sequences contribute to the genetic diversity of the human leukocyte antigen molecules. A map of the human MHC or HLA complex is shown in the Figure given below.
Class I Region
The class I region is subdivided into three major loci, termed HLA-A, HLA-B, and HLA-C, and minor loci consisting of HLA-G, HLA-E, HLA-F, HLA-H, and HLA-J and MHC class I chain-related genes (MIC). The major class I loci (A, B, and C) present antigens to CD8 T cells.
Minor loci play a variety of immunologic roles. HLA-E interacts with natural killer (NK) cell CD94 and NKG2 receptors to enhance or inhibit NK cell activity. HLA-F is a kind of HLA. an unidentified function HLA-G has a large number of self polypeptides. has a molecular structure that is identical to that of on the surface of extravillous cytotrophoblasts in high amounts as well as the placenta In pregnancy, HLA-G plays an important function. It’s unknown, but it could cause immunological responses to shifting from Th1 to Th2. HLA-H and HLA-J have unknown roles.
The MIC family is a non-traditional HLA gene complex. In the gut and synovia, gene products are produced as a stress response to virus infection or cellular injury. Their MIC gene products have a structure that resembles that of a class I molecule. Antigens are unable to be presented. The presence of MIC in a cell indicates that it is ready to divide. CD8 and NK cells destroy them.
Structure of class 1 molecules
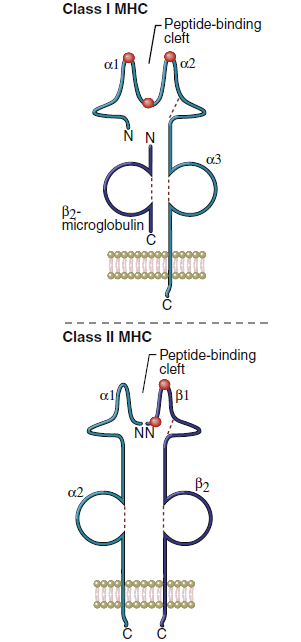
Class, I molecules comprise a single glycosylated α-chain (394 amino acids), which is noncovalently bound to β2- microglobulin (94 amino acids). The class I protein consists of two intracellular and three extracellular domains. The two intracellular domains attach the molecules to the cell membrane and extend into the cytoplasm. The three extracellular domains—designated α1, α2, and α3—are each composed of 90 amino acids. The α1- and α2- domains have unique helical structures that form an antigen-binding cleft used to present antigens to T cells.
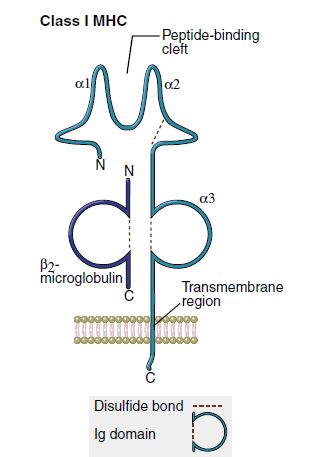
Peptides are tethered to the ends of the binding groove usually at positions 2 and 9. Thus, the binding cleft accommodates only small peptides containing 8 to 12 amino acids. Flexing of unbound peptides and side chains within the binding cleft creates a three-dimensional epitope structure that is recognized by T cells. The α3-domain stabilizes the interaction between class I molecules and the lymphocyte T cell receptor.
Class II Region
Class II gene products are induced and expressed on monocytes, macrophages, dendritic cells, and B cells. Within the class II region are three loci (DP, DR, and DQ) involved in antigen presentation. Class II molecules present antigens to CD4Th1 and CD4Th2 cells. Other genes in the class II region code for proteins that are important in antigen processing.
structure of class 2 molecules
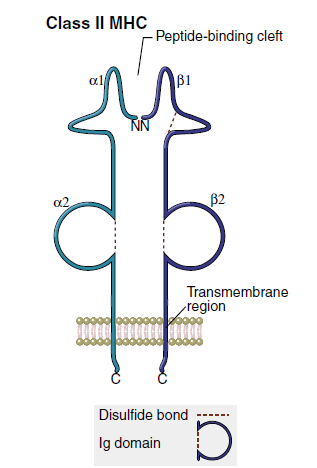
The class II region has three major loci (DP, DQ, and DR). Class II molecules consist of heavily glycosylated α- and β-chains. Although both chains have a similar structure, the α-chain is larger (30–34 kiloDaltons [kDa]) compared with the β-chain (26–29 kDa). An antigen-binding cleft comprises the α1- and β1-domains (Figure 4-3). The peptide binding to the class II molecule occurs in the middle of the binding cleft at positions 1, 4, 6, and 9. Peptides are bound in a manner analogous to a long pipe being held in a centrally located vise. Peptides are held in place by hydrogen bonding and van der Waals forces. All class II antigens bind at the same anchor positions. However, they are much larger than class I antigens (10–30 amino acids) and protrude from either end of the cleft.
Class III Region
The class III region has 62 genes, which have diverse functions. Most class III gene products are not membrane proteins but are secreted into the environment. Products include three complement components and the tumor necrosis factor (TNF).
Inheritance of human leukocyte antigens
A haplotype is a collection of HLAs (HLA A, B, C, DP, DQ, and DR) on the same chromosome. Each person’s genotype is made up of two haplotypes. N=6 HLA genes in one set are inherited from the father, and the other haplotype (N=6) is inherited from the mother.is a trait that is passed down from the mother. Both haplotypes are equally prevalent. and on the surface of cells, HLA genes are frequently passed down through the generations. as haplotypes in blocks, Deoxyribonucleic acid (DNA) is a type of nucleic acid that is found in the human body. Gene conversion is the process of exchanging (DNA) segments. During meiosis, two homologous chromosomes with identical sequences are produced. Multiple genes are mismatched when they are organized in tandem. Crossing over parental haplotypes and DNA recombination allows individual genes or parts of genes to transfer from one chromosome to another without any loss of function. The exchange of DNA causes multiple amino acid changes in the original gene and the formation of a recombinant HLA molecule.
heterozygosity
The inheritance of multiple genes at the same locus could be the outcome of a population balancing selection process, which theoretically gives heterozygotes a survival advantage. Although all HLA molecules can present antigens to T cells, they do so in different ways. The HLA molecule binds to a variety of antigens. The inheritance of paternal and maternal haplotypes is effectively doubled. enhances the host’s antigen-presenting capabilities and The likelihood that the majority of individuals in a species are heterozygotes. HLA heterozygotes are more resistant than homozygotes. Homozygotes are more susceptible to disease than homozygotes since they have a higher number of genes. HLA molecules that deliver antigens have a diverse repertoire.
Allelic polymorphism
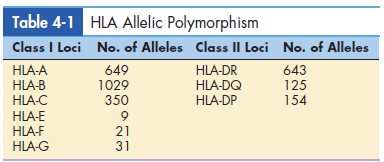
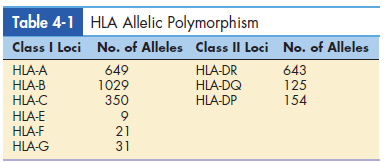
Allelic and single nucleotide polymorphisms give additional genetic variation that is important for disease resistance (SNPs). When there are numerous different versions of the same gene, it’s called polymorphism. The gene is polymorphic at the moment. Each iteration of An allele is a term for a polymorphic gene. There are more than 3000 different alleles. Within the HLA complex, are present. There are 2,000 polymorphic Only alleles in class I loci that have been reported. There are over 900 of them. Different alleles can also be identified in the DR, DQ, and DP loci of class II. HLA-F, HLA-G, HLA-H, and other nonclassical minor loci When compared to antigen-presenting cells, HLA-J is less polymorphic. locations and their tissue distribution are constrained.
Table 4-1 lists the different alleles found in major loci. Single base-pair alterations in genes that code for each HLA allele are known as SNPs. SNPs have the effect of generating new allele variants.
Nonsynchronous amino acid substitutions in the-helical sides of the binding cleft or changes in the peptide anchoring strands are linked to allelic polymorphism and SNPs. The cleft’s floor is formed by this. A modification to the binding cleft antigens attaches to the allele in a different fashion (Figure 4-4).
Allelic polymorphism is a response to the constant and The continuing evolution of microbes is essential to the survival of the species. Within an HLA locus, the presence of Different alleles presents a wide range of antigens from The same microbe ensures that some members of a species will survive the microbial infection. In addition, allelic polymorphism A microbe protects some members of a species when a inadvertently expresses a specific HLA allele. Microbes that express an HLA allele would be considered “self ” and evade immune detection. As a consequence of infection, significant population mortality may occur among individuals carrying the allele. However, individuals expressing other HLA polymorphic alleles within the same loci would be unaffected.
human leukocyte antigen single
Nucleotide polymorphisms and disease
- The HLA molecule’s ability or inability to present antigens to T cells determines an individual’s resistance or susceptibility to microbial infections.
- For example, self-limiting hepatitis B is associated with the expression of HLA-DR13.
- Individuals homozygous for DR13 have more efficient presentation of hepatitis antigens, a strong and vigorous CD4 response, and accelerated viral clearance.
- Chronic hepatitis B infections develop in individuals expressing other HLA DR molecules.
- HLA alleles also control susceptibility to infections.
- The strongest relationship between HLA and disease is found in HLA alleles containing SNPs.
- For example, individuals expressing (DRB1*1501) molecules have a high risk of developing pulmonary tuberculosis.
- It is assumed that these class II molecules can not present mycobacterium antigens to CD4Th1 cells.
Antigen-presenting molecules outside the human leucocyte antigen complex
- The CD1 family of genes is involved in antigen presentation.
- Unlike HLA molecules, they are encoded by a gene clusteron chromosome 1.
- The configuration of the CD1 molecule is similar to a class I protein with respect to subunit organization and the presence of B2-microglobulin.
- CD1 molecules present highly conserved glycolipids to CD4Th1 cells.
- Glycolipids include mycolic acid, glucose monomycolate, phosphoinositol mannosides, and lipoarabinomannan derived from the Mycobacterium species that cause tuberculosis and leprosy.
- Two families of CD1 molecules exist: (1) Group I consists of CD1a, CD1b, and CD1c. (2) CD1d is the only member of group II (Table 4-2).
- CD1a is expressed on thymocytes, dendritic cells, and Langerhans cells.
- CD1b is expressed on monocytes and macrophages.
- CD1c is found on circulating B cells, the splenic mantle zone, and tonsillar B cells.
- It is unclear whether the five members of the CD1 family present the same antigens or different antigens.
- It is conceivable that each represents a redundant system for the identification of conserved molecules.
- Some evidence suggests that group I CD1 is promiscuous and presents an overlapping set of glycopeptides.
- Other CD1 molecules may have individual binding properties.
Antigen-presenting molecules and immunodeficiency
Bare Lymphocyte Syndrome
The bare lymphocyte syndrome (BLS) is a form of severe combined immunodeficiency (SCID). In this syndrome, individuals B cells lack class II molecules on the surface of B cells. monocytes Class II molecules are not produced because of the mutations in the consensus sequences of gene promoters that regulate HLA class II structural genes. Mutations prevent the docking of regulator factor X (RFX) and class II transactivator (CIIT) with the promoter genes. CIIT regulates the expression of the class II molecule. Children with BLS have repeated infections with Candida albicans or Pneumocystis jiroveci. Common childhood viral infections with a respiratory syncytial virus (RSV) or cytomegalovirus (CMV) are invariably fatal.
Treatment of Bare Lymphocyte Syndrome
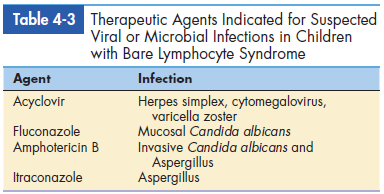
Infants with this disease die within the first year of life unless they receive hematopoietic stem cell transplantation. For suspected microbial and viral illnesses, aggressive antibiotic therapy is recommended. The agents used to treat BLS are listed in Table 4-3.